Honlapunkon már többször leírtuk, milyen gyönge lábakon áll a víruskimutatás, és mennyire logikátlan a citopátiás hatás (CPE) elfogadása bizonyítékképp a vírus létezésére [1]. A mai alkalommal Mike Stone írása alapján részletesen bemutatjuk, miért nem fogadható el a vírusok létének bizonyítékaképp a citopátiás hatás.
| Kivonat (a bejegyzés terjedelmére tekintettel)
A szerző bemutatja, milyen okok zárják ki, hogy a citopátis hatás a vírusok létezését bizonyítja. „Szeretném előrebocsátani, hogy a sejtkultúra-kísérlet többszörösen is áltudományos tákolmánynak bizonyult. Már írtam a kísérlet tervezésébe beépített logikai hibákról, és bemutattam, hogy az miért nem felel meg a tudományos módszernek. Alapvető: a kísérlet nem egy megfigyelt természeti jelenségből származó hipotézist tesztel. A kísérlet előtt nincs érvényes független változó – nincsenek tisztított, izolált „vírusos” részecskék. A függő változó, a citopátiás hatás (CPE) egy nem specifikus, laboratóriumban előidézett produktum, amely több okkal is magyarázható anélkül, hogy egy kitalált okra lenne szükség. Ennek ellenére az a állítás, hogy a CPE egyenlő a „reprodukáló vírus” jelenlétével, a virológia alapjává vált. Célom itt annak bemutatása, hogy ez az alapvető feltételezés soha nem lett bebizonyítva, kezdettől fogva alaptalan volt, és a virológia saját kísérletei és módszerei cáfolták.” A laboratóriumban in vitro körülmények között létrehozott morfológiai változásokat és végül a sejtek elhalását John Franklin Enders virológus az 1940-es évek végén citopátiás hatásoknak (CPE) nevezte el. Ezeket a változásokat hamarosan a „vírusreplikáció” bizonyítékaként értelmezték. Ezek a kísérletek soha nem tartalmaztak tisztított és izolált „vírus” részecskéket. Ehelyett a „poliovírust” állítólag tartalmazó szöveti homogenizátumokat adtak emberi embrionális szövetek tenyészetéhez, és a későbbi sejtpusztulást „vírus szaporodásnak” értelmezték. Ezt a ugrást – a korrelációtól az ok-okozati összefüggésig – soha nem igazolták empirikusan. Az ugrás a feltételes megfigyelésről a biztosra vett állításra – a modern virológia alapjává vált. Enders „felfedezéséért” 1954-ben Nobel-díjat kapott. De még abban az évben közzétett egy tanulmányt, amelyben elismerte, hogy a központi bizonyíték – a citopátiás hatások (CPE) – nem specifikusak a „vírusokra”. Ackermann amerikai biológus szintén 1954-ben arra a következtetésre jutott, hogy a „vírusreplikáció” és a sejtkárosodás elkülöníthető jelenségek, azaz a CPE nem függ szigorúan a „vírus szaporodásától”. A folyamatokat „jelentős fokú autonómiával” jellemezte. Kísérletei arra utaltak, hogy bármi is okozza a sejtkárosodást (amit a virológusok „vírusos citopatogenitásnak” értelmeztek), az nem egyszerűen azonos a „vírusreplikációval”. Ez az eredmény aláásta azt a feltételezést, hogy a tenyészetben bekövetkező sejthalál = „vírusreplikáció” = „vírusaktivitás” bizonyítéka. Ackermann eredményei messze nem bizonyították a „vírusreplikációt”, hanem a tenyésztésen alapuló vizsgálatok kétértelműségét tárták fel, és arra utaltak, hogy amit a virológusok „citopatogenitásnak” neveztek, gyakran a kísérleti rendszerük nem specifikus toxikus hatása volt. Munkája a virológia saját módszereinek és szabványainak felhasználásával kimutatta, hogy a CPE nem megbízható mutatója a „vírusreplikációnak”. Ehelyett ugyanolyan könnyen magyarázható a sejtek folyamatainak zavarával és a tenyésztési rendszer toxikus melléktermékeivel. Kritikus megállapításai – különösen a toxin által kiváltott citopátiás hatás azonosítása – tartós hatással voltak arra, ahogyan a virológusok értelmezik a tenyésztésen alapuló vizsgálatokat és a „vírusfertőzés” biokémiai alapjait. Döntő fontosságú, hogy feltárták, mennyire nem specifikus és félrevezető volt valójában a tenyésztésen alapuló vizsgálat. Ennek hatására a virológia egy sor értelmező „mentőeszközt” fejlesztett ki a domináns hipotézis megőrzése érdekében. H.G. Pereira virológus szintén megerősítette, hogy CPE jelentkezik vírusanyagok kimutatható termelődése nélkül, és fordítva, vírusmultiplikáció jelentkezik nyilvánvaló CPE nélkül. Dokumentálta továbbá, hogy a tenyésztési körülmények megváltoztatásával indukálható vagy elnyomható volt a CPE. Míg kezdetben (Enders óta) a CPE-nek nevezett morfológiai változások és sejtelhalás voltak a bizonyítékok a vírusok szaporodására, később az ellentmondásos eredmények és esetenként a CPE hiányának hatására egyéb jelenségeket is elfogadtak a vírusok szaporodásának bizonyítékaként, úgy mint a hemadszorpciót, az interferenciát, az antigénfestést vagy a genetikai fragmenseket. Tehát: Ha CPE ⇒ akkor vírus |
A szövetkultúra-technikák megjelenése a 20. század elején fordulópontot jelentett a kezdő virológia területén, lehetővé téve a kutatók számára, hogy laboratóriumban olyan hatásokat hozzanak létre, amelyeket aztán láthatatlan entitásaiknak tulajdoníthattak. A szövet- és sejtkultúra-technikák megjelenése előtt a „vírusok” csupán egy változó fogalom voltak, amelyet kizárólag negatív értelemben határoztak meg. Ahogy Thomas Rivers megjegyezte, azokat kizárásos alapon azonosították:
- a hagyományos mikroszkóppal NEM láthatók,
- szűréssel NEM nyerhetők ki,
- élő sejtek nélkül NEM képesek szaporodni.
A szövetkultúra 20. század eleji megjelenésével a virológia végre nyert valami láthatót – de csak a sejtkárosodás formájában. Fontos azonban megjegyezni, hogy a szövet- és sejtkultúrák nem természetes biológiai rendszerek. Mesterséges laboratóriumi konstrukciók, amelyek haldokló vagy legyengült állati és emberi szövetekből készülnek, és táptalajok, borjú szérum, antibiotikumok, gombaellenes szerek és különböző kémiai adalékok segítségével tartják őket életben. Ezek a keverékek semmiben sem hasonlítanak egy élő szervezet belső környezetére. Az ilyen rendszerekben megfigyelt hatások a kultúra körülményeinek lebomlását, toxicitását és stresszét tükrözik, nem pedig egy természetes folyamatot. Más szavakkal, a tenyésztés nem a természetben megfigyelt jelenség és annak okának kutatásából származik, hanem először a jelenséget hozták létre, és csak utána feltételezték az okát.
A laboratóriumban létrehozott jelenségek közül a legszembetűnőbbek a morfológiai változások voltak, mint például a kerekedés, az elválás, a szincítiumok (több sejtmagot tartalmazó sejtegység, sejtfal nélkül egymásba folyó sejtek) kialakulása és végül a sejtek elhalása – ezeket a változásokat John Franklin Enders virológus az 1940-es évek végén citopátiás hatásoknak (CPE) nevezte el. Ezeket a változásokat hamarosan a „vírusreplikáció” bizonyítékaként értelmezték.
Thomas Weller virológus szerint – aki Endersszel együtt dolgozott a polio kutatásokban – az úgynevezett „vírusok” degeneratív változásokat idéztek elő azokban a sejtekben, amelyekben szaporodtak, és ezek a változások morfológiailag vagy biokémiailag kimutathatók voltak. Ezeket a látható változásokat kezdettől fogva a „vírusos” replikáció közvetlen bizonyítékaként értelmezték, ami egy központi feltételezést alapozott meg: a „vírus” megfertőzi a sejtet, replikálódik, és ez a replikáció CPE-t eredményez. Ez a nem tesztelt feltételezés gyorsan alapjává vált a „vírusizolálásnak”, az azonosításnak és a mennyiségi meghatározásnak. Az Encyclopedia of Virology a CPE-t úgy definiálta, mint amikor „a vírusok megölik vagy morfológiailag módosítják gazdasejtjeiket, amikor szaporodnak”. Hasonlóképpen, a Fields Virology megjegyzi, hogy „a vírusok sejtekben történő replikációja kimutatásának egyik klasszikus módja a vírusfertőzés eredményeként bekövetkező sejtszerkezeti változások, vagyis a CPE-k megfigyelése volt”.
Az 1980-as évek végén még szilárdan tartotta magát ez a feltételezés. A Veterinary Virology című könyvben Frank Fenner a CPE-t „a sejtkárosodás látható bizonyítékaként” írta le, amely akkor következik be, amikor a „vírusok” megölik azokat a sejteket, amelyekben replikálódnak:
„Sok vírus megöli azokat a sejteket, amelyekben replikálódik, így a fertőzött sejtek monorétegeiben fokozatosan megjelennek ennek a sejtkárosodásnak a látható jelei, ahogy az újonnan kialakult virionok terjednek, és a tenyészet egyre több sejtjét vonják be a folyamatba. Ezeket a változásokat citopátiás hatásoknak (CPE) nevezik, és a felelős vírust citopatogénnek nevezik.”
Hasonlóképpen, az Altmeyer’s Encyclopedia – egy orvosi szakemberek számára készült referencia – a CPE-t „a vírus replikációjának a gazdasejtre gyakorolt hatásaként” definiálja.
Összességében ezek a definíciók mutatják, milyen mélyen gyökerezik a CPE = „vírusreplikáció” feltételezés. A CPE megfigyelését már nem magyarázatra szoruló jelenségként kezelték, hanem inkább a láthatatlan entitás jelenlétének és szaporodásának közvetett jelzőjeként.
A kritikus kérdés azonban továbbra is megválaszolatlan maradt: ez a laboratóriumban létrehozott jelenség valóban bizonyítja-e, hogy „vírusok” voltak jelen és replikálódtak, vagy csak feltételezték ezt? Ha más bizonyítékok kerültek volna napvilágra, amelyek cáfolták volna a CPE = replikáció dogmát, akkor a virológia keretei egyáltalán lehetővé tették volna ennek elismerését?
Járjuk körbe a témát.
Klasszikus bizonyítás: Enders és a „vírusos” replikáció „bizonyítéka”
Enders, Weller és Robbins 1940-es évek végi munkája példázza: a „vírusreplikáció” bizonyítás számára egzisztenciálisan fontos volt a támaszkodás a CPE-re.
Grafe „A kísérleti virológia története” című műve (1994, 4. fejezet) szerint Enders csoportjának célja a „poliovírus” „vírusos szaporodásának” bizonyítása volt. Megfigyelték, hogy az állítólag „poliovírust” tartalmazó minták, amelyeket emberi embrionális szövetbe oltottak be, morfológiai változásokat okoztak, amelyeket specifikus „immun” szérum gátolt:
„Enders és kollégái 1949-ben egy nagyon jelentős fejlesztést vezettek be, amely döntő fontosságú volt a kvantitatív virológia számára. F. Robbins 1948-ban szövetkultúra-kísérletekben bizonyította, hogy az egérhez adaptált Lansing poliovírus-törzs szaporodik az emberi embrionális bélszövet kultúrájában. Hasonló eljárással a vírust sikeresen tenyésztették emberi embrionális ideg-, bőr-, kötőszövet- és izomszövetben is. A vírus szaporodásának bizonyítékát későbbi egerekkel és majmokkal végzett fertőzési kísérletek során kapták meg. Ennél azonban sokkal fontosabbak voltak a sejtek morfológiájában bekövetkezett változások, valamint a fertőzött sejtmagok festésképtelenségének felfedezése. Egy 1950-es tanulmányban ez a két tudós Wellerrel együtt bemutatta a „citopátiás hatásokat”, amelyeket később a citopátiás vírusok citopátiás hatásának (CPE) neveztek el.
„A bizonyíték, hogy a poliovírus, amelyet addig szigorúan neurotróp vírusként osztályoztak, in vitro szaporodhat nem idegszövetben, más típusú vírusokkal végzett hasonló szövetkultúra-kísérleteket indított el. Meg kell említeni, hogy az Enders-csoport célja a poliovírus szaporodásának igazolása volt a bélrendszerben.”
Döntő fontosságú azonban, hogy ezek a kísérletek soha nem tartalmaztak tisztított és izolált „vírus” részecskéket. Ehelyett a „poliovírust” állítólag tartalmazó szöveti homogenizátumokat adtak emberi embrionális szövetek tenyészetéhez, és a későbbi sejtpusztulást „vírus szaporodásnak” értelmezték. Ezt a ugrást – a korrelációtól az ok-okozati összefüggésig – soha nem igazolták empirikusan.
Ezeket a megfigyeléseket mindazonáltal úgy értelmezték, hogy megerősítik, hogy a feltételezett „vírus” okozta a megfigyelt CPE-t, és tovább használták a „vírus” tartalom számszerűsítésére, a „vírusos” törzsek azonosítására és az „antivirális” anyagok tesztelésére. A trió 1954-ben Nobel-díjat kapott a polio-kutatásai alapján, a díj indoklása szerint „a poliomyelitis vírusok különböző típusú szövetkultúrákban való szaporodási képességének felfedezéséért”. A Nobel-bizottság megfogalmazása egyértelművé teszi, hogy a kutatók megfigyeléseinek (a tenyésztett sejtekben megfigyelt CPE) értelmezését „vírusos növekedés/replikáció” bizonyítékaként fogadták el. Azáltal, hogy ezt az értelmezést „felfedezésként” rögzítették, a díj gyakorlatilag eltörölte a feltételezés és a bizonyíték közötti különbséget, megerősítve ezzel azt a feltételezést, hogy a CPE egyet jelent a „vírusos” növekedéssel. Ettől a pillanattól kezdve a virológia olyan keretek között folyt, ahol a tenyészetben látható sejtkárosodást a „vírusreplikáció” egyértelmű bizonyítékaként kezelték.
De valóban ezt figyelték meg Enders és társai?
Enders kezdeti óvatossága és ellentmondásai
1949-ben megjelent „A Lansing-törzsű poliomyelitis vírus tenyésztése különböző emberi embrionális szövetek kultúráiban” című cikkükben Enders hosszú távú tenyésztés után morfológiai különbségeket észlelt az oltott és az oltatlan kultúrák között: az oltott fragmensek sejtjeinek magjai jelentős festődési tulajdonságvesztést mutattak, míg a kontrollkultúrák sejtjei normálisnak tűntek. Enders azonban nem sietett elhamarkodott következtetéseket levonni, hanem így írt:
„Mivel a vizsgált anyag mennyisége még viszonylag kicsi, nem lehet azt a következtetést levonni, hogy az oltott kultúrákban megfigyelt változásokat a vírus okozta.”
Ez a beismerés figyelemre méltó. Enders nyíltan elismerte, hogy a korreláció nem bizonyítja az ok-okozati összefüggést, és hogy a megfigyelt sejteltéréseknek lehetnek más magyarázatai is. Ebben a korai szakaszban a CPE és a „vírusreplikáció” közötti kapcsolatot nem tényként kezelték, hanem további megerősítést váró munkahipotézisnek.
De alig egy évvel később, az 1950-es, „A poliomyelitis vírusok in vitro citopatogén hatása emberi embrionális szövetekre” című cikkben Enders és kollégái teljes fordulatot vettek, és arra a következtetésre jutottak, hogy a megfigyelt citopatogén hatások „nem hagynak kétséget afelől”, hogy a poliomyelitis „vírus” szaporodik a sejtekben, és hogy a CPE megbízható marker lehet a „vírusok” azonosítására. Ez a ugrás – a feltételes megfigyelésről a biztosra vett állításra – a modern virológia alapjává vált:
„Ezek a jelenségek két általános szempontból érdekesek. Először is, nem hagynak kétséget afelől, hogy a poliomyelitis vírus in vitro szaporodhat az idegrendszer sejtjein kívül más sejtekben is, és azokban súlyos károsodást okozhat. Másodszor, olyan kritériumokat nyújtanak, amelyek alapján a vírus jelenléte in vitro felismerhető, és így alapot adhatnak a vírus betegekből vagy állatokból való izolálásának, a vírus mennyiségi vizsgálatának, a szerológiai tipizálásnak és esetleg a kemoterápiás és antibiotikumos anyagok szűrésének technikájához. További tanulmányok szükségesek ezeknek a jelenségeknek az ilyen célokra való alkalmazásának megbízhatóságáról és megvalósíthatóságáról.”
Azonban csak néhány évvel később, ugyanabban az évben, amikor Nobel-díjat kapott azért, hogy „bizonyította”, hogy a „vírusok szaporodnak” a szövetkultúrákban, és közzétette az érdeklődés központjába került kanyaró-tanulmányát, amely szintén CPE-t mutatott ki a „nem fertőzött” kultúrákban, Enders egy informatív cikket tett közzé Cytopathology of Virus Infections: Particular Reference to Tissue Culture Studies (Vírusfertőzések citopatológiája: különös tekintettel a szövetkultúra-vizsgálatokra) címmel. Ebben Enders elismerte, hogy a központi bizonyíték – a citopátiás hatások (CPE) – nem specifikusak a „vírusokra”. Elismerte, hogy
- a CPE-t „sok káros anyag” okozhatja, nem csak a „vírusok”.
- Az inklúziós testek [2] nem bizonyító erejűek, mivel vegyi anyagok és ismeretlen tényezők is előidézhetik őket.
- A tenyésztési körülmények és a donorok változói befolyásolják az eredményeket, vagyis a szövet korának, fajának és előzetes kezelésének függvénye lehet, hogy CPE jelentkezik-e.
- A „vírusos” okokra vonatkozó következtetések csak „feltételesek” voltak.
Enders így írt:
„A fenti 1. csoportban említett jelenségek számos károsító ágens által kiválthatók. Ennek megfelelően önmagukban nem tekinthetők feltétlenül vírusaktivitás eredményének. Ennek bizonyításához bizonyos ellenőrzési eljárásokat (sorozatos tenyésztés, homológ antitestekkel történő változások megelőzése stb.) kell alkalmazni. Azonban egy adott vírus hatásának ismerete egy adott sejtrendszerben gyakran lehetővé teszi a megfigyelő számára, hogy ideiglenesen arra a következtetésre jusson, hogy ez a vírus felelős a jelenségért.”
„A vírusos károsodás morfológiai mutatói közül a legjellemzőbb a zárványtestek kialakulása (fenti 2. csoport), bár ez a folyamat sem tekinthető a vírusaktivitás döntő bizonyítékának, mivel bizonyos kémiai és egyéb ismeretlen tényezők is befolyásolhatják kialakulását. A zárványtestek voltak az első citopátiás változások, amelyeket in vitro kerestek és a fertőzés kritériumaként alkalmaztak. A vírus szaporodásának mutatóiként azonban kevésbé hasznosak, mint az 1. csoport változásai, mivel ezek a struktúrák csak festett preparátumokban mutathatók ki egyértelműen.”
„Az in vitro citopatogenitást olyan tényezők befolyásolják, amelyek közül néhány ismert, míg sok még meghatározásra vár. Bevezetésként néhány, jelenleg ismert tényezőt említünk, majd áttekintjük az egyes kórokozók viselkedésével kapcsolatos rögzített megfigyeléseket. Elsődleges fontosságú az a faj, amelyből a sejtek származnak. A vírus gazdaszervezet-tartományához hasonlóan a tenyésztett sejtekben is megfigyelhető a citopatogenitás tartománya. Azonban az organizmus és sejtjei in vivo érzékenysége között nem mindig áll fenn összefüggés. Bár ez az összefüggés gyakran fennáll, az érzékeny fajok szövetei néha nem támogatják a vírus szaporodását, míg ennek ellenkezője is előfordul.
A szövet donorának életkora befolyásolhatja a citopatogenitást. Ahogyan a fiatal állatok gyakran fogékonyabbak a fertőzésekre, úgy szövetük is érzékenyebb lehet a vírus által okozott károsodásokra, azonban ez az összefüggés sem állandó. A legtöbb releváns adat arra utal, hogy a vírusfertőzésekkel szemben szerzett immunitás nem tükröződik a sejtek fokozott ellenállóképességében, ami technikai szempontból előnyös, mivel így nem kell aggódni a donor állat immunológiai állapotáért.
A citopátiás károsodás intenzitása és mértéke a vírus törzsétől vagy a szövetkultúrában történő vizsgálatát megelőzően történő szaporodási körülményektől függően változhat. A kutatónak fel kell készülnie arra, hogy egy vírusfaj számos képviselőjének vizsgálata során ilyen eltérésekkel találkozik. A mérsékelt vagy gyenge citopátiás hatást néha fokozhatja a sorozatos in vitro átvitel.
Más szavakkal, pontosan abban a pillanatban, amikor a virológiát a „vírusreplikáció” bizonyításával emelték magasabb szintre, annak vezető alakja nyíltan elismerte, hogy a bizonyítékok kétértelműek, zavarosak, és feltételezéseken alapulnak.
Két évvel később, 1956-ban azonban Enders másképp fogalmazott. Gordon Wilson előadásában, amelynek címe „Megfigyelések bizonyos, emberben exantémás (bőrkiütéseket okozó betegségek összefoglaló neve) betegségeket okozó vírusokról” volt, elmagyarázta, hogy a szövetkultúra „hiányosságait” nem a kórokozó izolálásával és független bizonyításával sikerült leküzdeni, hanem a gyanús anyaggal való érintkezés után a sejtekben bekövetkező degeneratív változások megfigyelésével:
„Ebben a tekintetben a szövetkultúra hiányosságait sok esetben sikerült leküzdeni azzal, hogy bebizonyították, hogy a eddig vizsgált vírusok többsége szaporodásakor degeneratív változásokat okoz a sejtekben. Ezek a változások gyakran már néhány órával vagy néhány nappal a vírusinokulum bejuttatása után láthatóvá válnak, és gyakran elég jellegzetesek ahhoz, hogy lehetővé tegyék a vizsgált vírus ideiglenes azonosítását. Ezeket a hatásokat ma „citopátiás változásoknak” vagy „a vírus citopatogén hatásainak” nevezik.
Bár a citopátiás változásokat már korábban is észlelték különböző kutatók, különösen Ivanovics és Hyde 1932-ben, valamint Huang 1942-ben, teljes jelentőségüket csak 1950-ben ismerték fel, amikor bebizonyosodott, hogy a poliomyelitis vírusai számos emberi szövet kultúrájában erős citopátiás hatást mutat. Azóta számos vírussal és különböző fajokból származó sejtekkel elért eredmények azt mutatják, hogy a citopátiás hatások in vitro megjelenése ugyanolyan megbízható kritériumot jelent a vírusok szaporodására nézve, mint az állatokban jelentkező fertőzés tünetei. Ráadásul ezek a kritériumok gyakran kényelmesebben és pontosabban megfigyelhetők.
Bár Enders korábban elismerte, hogy a CPE nem specifikus és több tényező is okozhatja, most a mesterséges sejtrendszerekben megfigyelt vizuális degenerációt bizonyítékként emelte ki. A megfigyelt sejthalált „a vírus szaporodásának olyan megbízható kritériumaként” üdvözölte, „mint az állatokban jelentkező fertőzés tüneteinek megjelenése”. Ezeket a laboratóriumi artefaktumokat a betegségnek megfelelően kezelték, anélkül, hogy előbb függetlenül igazolták volna az okozó kórokozót.
Annak ellenére, hogy a CPE-t a „vírusok szaporodásának” megbízható mutatójaként írta le, egy évvel később, 1957-ben megjelent Comments on Viral Cytopathogenicity (Megjegyzések a vírusok citopatogenitásáról) című cikkében Enders hangsúlyozta, hogy valójában milyen keveset tudnak a „vírusok citopatogenitásának” feltételezett mechanizmusáról. Nyíltan elismerte, hogy az alapul szolgáló folyamat teljesen rejtélyes maradt, és hogy egyik hipotézis sem volt kielégítő – különösen azért, mert egyik sem tudta megmagyarázni, hogyan történhet „vírusreplikáció” sejtkárosodás (CPE) nélkül. A magyarázatot a jövő kutatóira bízta:
„A vírusok citopatogenitásának problémájáról szóló töredékes megjegyzéseimet azzal a megállapítással szeretném befejezni, hogy hangsúlyozom, mennyire keveset tudunk az alapjául szolgáló mechanizmusokról. Jelenleg egyszerűen nem tudjuk, hogy ezek az egyedülálló biológiai entitások hogyan indítják el azt a bonyolult intracelluláris eseménysorozatot, amely az imént leírt hatásokhoz vezet. Különböző hipotézisek születtek. Számomra egyik sem teljesen kielégítő, mivel nem veszik figyelembe a két legkiemelkedőbb jellemzőt, amit felsoroltam, azaz a különböző vírusok által kiváltott hatások sokféleségét és azt, hogy bizonyos körülmények között a vírusok szaporodnak anélkül, hogy látható sejtkárosodást okoznának. De bízunk benne, hogy a biokémikusok és biológusok közös erőfeszítései végül teljes képet adnak a vírusos betegségekért felelős tényezőkről. Megkockáztatom azt a jóslatot, hogy a kielégítő magyarázat nem lesz sem egyszerű, sem hamarosan elérhető.”
Más szavakkal, még évekkel azután is, hogy állítólag bizonyította a „vírusreplikációt”, Enders elismerte, hogy valójában senki sem tudta, mi okozza a megfigyelt sejt hatásokat. A feltételezett „vírusmechanizmus” hipotetikus maradt, és azok a jelenségek, amelyekért Nobel-díjat kapott, továbbra is megmagyarázhatatlanok maradtak.
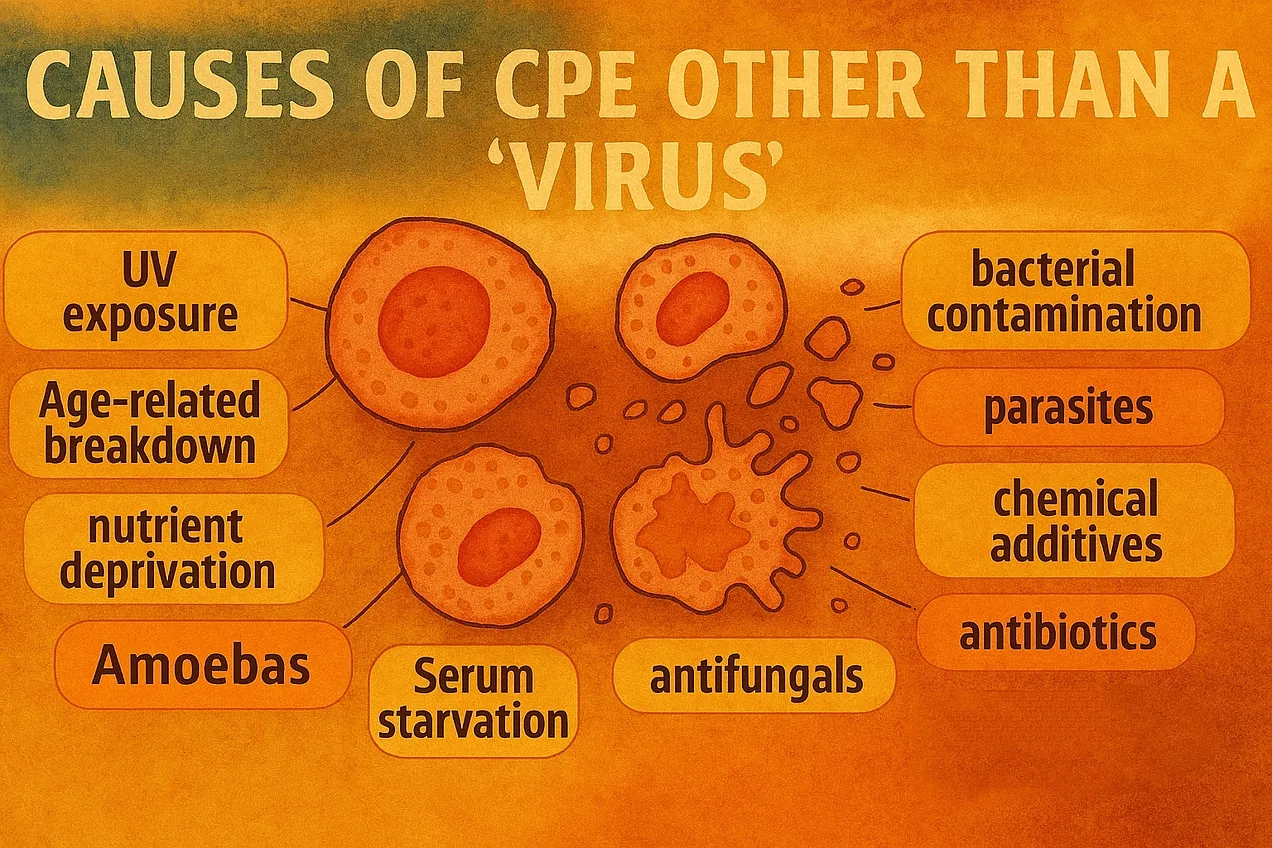
Egyéb, CPE-t kiváltó okok a víruson kívül: UV-sugárzás, öregedés, tápanyag-megvonás, amőbák, szérum-megvonás, fungicidek, antibiotikumok, vegyianyagok, paraziták, bakteriális fertőzés
Visszatekintve Enders maga is leírta 1972-ben, hogyan jutottak ezekre a következtetésekre. Korai szövetkultúra-kísérleteire visszagondolva elmagyarázta, hogyan keresték a „látható bizonyítékot” arra, hogy „vírus” van jelen a sejtkultúrában. Lényegében feltételezték, hogy az eredeti mintában „vírus” volt, majd azt állították, hogy a „fertőzött” és a „nem fertőzött” kultúrák közötti bármilyen különbség bizonyítja annak jelenlétét:
„1948–1949-ben Weller, Robbins és én azt találtuk, hogy a poliovírusok in vitro képesek szaporodni az emberi szövetekben az idegszöveteken kívül is. Ez a felfedezés akkoriban különösen érdekes volt, mivel a legtöbb ember akkoriban úgy vélte, hogy ezek a kórokozók szigorúan neurotrópikusak és más típusú sejtekben nem képesek szaporodni. Sokak számára a kísérleteink legjelentősebb eredménye az volt, hogy bebizonyítottuk, hogy ez a nézet téves. Azonban szinte azonnal rájöttünk, hogy bár megfigyeléseink hozzájárulhatnak a poliomyelitis patogenezisének megértéséhez, gyakorlati alkalmazásuk a diagnózisban és a megelőzésben csak korlátozott lenne, ha egyáltalán lenne, hacsak nem találunk látható bizonyítékot a vírus jelenlétére magában a tenyészetben. Ezért nagyon korán elkezdtük keresni a vírusnak kitett sejtekben bekövetkezett változásokat. Először Maitland-típusú tenyészetet használtunk – csak apróra vágott embrionális szövetdarabokat. Ezeket rögzítettük, metszeteket készítettünk belőlük, és a vírus beoltása után különböző időpontokban megvizsgáltuk őket, összehasonlítva a beoltatlan tenyészetekből származó darabokkal. Találtunk olyan jeleket, amelyek arra utaltak, hogy a vírus romboló hatást gyakorolt a sejtekre. További munkát végeztünk ezen a döntő ponton, megerősítve a Maitland-tenyészetekben tett megfigyeléseinket, majd folytattuk az ilyen darabok túlélésének és a sejtek növekedésének tanulmányozását, miután a szövetet eltávolítottuk a lombikból és plazma cseppekbe helyeztük. Végül a hengeres csőben létrehozott tenyészetek egyrétegű növekedését tettük ki a vírusnak. Ez az eljárás bizonyult a legalkalmasabbnak, és nem hagyott kétséget a citopátiás hatásról, amely ilyen körülmények között gyors és specifikus volt, a továbbiakban pedig specifikus polio szérummal semlegesíthető volt, és felhasználható volt a vírus fertőzőképességi titerének meghatározására.
Ez a őszinte leírás feltárja Enders megközelítésének körkörös érvelését. Ő és kollégái azzal a feltételezéssel indultak ki, hogy a tenyészetekbe bejuttatott anyagban „vírus” van jelen. Ennek ellenére ebben a szakaszban még nem volt közvetlen bizonyítékuk arra, hogy ilyen entitás létezik a mintákban. Enders még azt is elismerte, hogy a szövetkultúra-kísérleteknek nincs gyakorlati jelentőségük, hacsak nem jelenik meg valamilyen látható jel, amely megerősíti feltételezésüket.
Más szavakkal, a hit megelőzte a bizonyítékot.
Amikor végül megjelent a sejtkárosodás – a citopátiás hatás –, azt a rég várt megerősítésként értelmezték, hogy a „vírus” végig jelen volt. De mivel előzetesen nem izoláltak egyértelmű, tisztított entitást, amely független változóként szolgálhatott volna, a CPE megjelenése logikailag nem igazolta feltételezésüket. A következtetés egyszerűen megismételte az előfeltevést: azzal kezdték, hogy feltételezték a „vírus” létezését, és azzal fejezték be, hogy ezt a feltételezést bizonyítottnak tekintették.
Enders azt is megjegyezte, hogy hasonló „citopátiás” megfigyelések már jóval az ő munkája előtt is léteztek, de vagy figyelmen kívül hagyták őket, vagy nem vonták le a megfelelő következtetéseket. A korai tanulmányokat gyakran akadályozták technikai korlátok, szennyeződés és erős értelmezési elfogultság. Nyíltan megkérdőjelezte, hogy miért nem vizsgálták alaposabban ezt a korábban megfigyelt, laboratóriumban létrehozott hatást, és két fő okot feltételezett:
- Az antibiotikumok hiánya, ami a szövetkultúra-kísérleteket rendkívül érzékennyé tette a szennyeződésre.
- A poliomyelitis drámai jellege, amely végül arra késztette a kutatókat, hogy komolyan vegyék figyelembe ezt a jelenséget.
Ez egy monumentális beismerés, mivel azt mutatja, hogy a mesterséges CPE „vírusbizonyítékká” emelésének motivációja nem empirikus bizonyítékon alapult, hanem gyakorlati akadályok és egy ijesztő betegség által kiváltott érzelmi és szociológiai nyomás kombinációján:
Nem tudom, miért nem használták ki az emberek olyan sokáig azt a tényt, hogy sok vírus in vitro citopatogén, mert ezt a tulajdonságot már többször is megfigyelték. Például Ivanovic és Hyde korai megfigyelései a Rivers III vírusról; Plotz és Ephrussi megfigyelései a baromfi pestis vírusról az 1930-as években; valamint Huang megfigyelései az 1940-es évek elején, aki gyönyörű megfigyeléseket tett a ló-encephalitis vírus citopatogenitásáról szövetkultúrákban. 1939 körül R. Relovával közösen, még nem publikált tanulmányokban ugyanazt a jelenséget figyeltük meg, azaz a csirkeembrió sejtjeinek teljes elpusztulását e kórokozó hatására. Tanulságosnak és kissé elgondolkodtatóan találom, hogy senki sem aknázta ki teljes mértékben e jelenség potenciális értékét. Úgy vélem, csak a poliomyelitis drámai jellege miatt vált általánosan elismertté jelentőségének, és tárták fel széles körben lehetőségeit. Kérdéses, hogy miért nem folytatták a korai megfigyeléseket. Talán az egyik ok az volt, hogy nem álltak rendelkezésre antibiotikumok. Így nem lehetett beoltani és tanulmányozni a fertőzött anyagokat szövetkultúrában. Ráadásul a tenyésztési technikák viszonylag fejletlenek voltak, és ennek eredményeként a megállapítások gyakran nem voltak kielégítően konzisztensek. Mindig is úgy éreztem, hogy Carrel nagy hatása a területen bizonyos értelemben korlátozó volt, mert a bakteriális fertőzés elkerüléséhez elengedhetetlennek tartott bonyolult óvintézkedések sokakat elriasztottak, akik egyébként alkalmazták volna a módszert. De bármi is legyen az oka, mint mindannyian tudjuk, néha hosszú időbe telik, mire a lényegileg fontos megfigyeléseket általánosan elismerik és alkalmazzák.”
Enders antibiotikumokkal kapcsolatos megjegyzése nem elhanyagolható. Valójában 1954-es Nobel-díj átvételekor tartott beszédében kijelentette, hogy az antibiotikumok felfedezése „forradalmat hozott a szövetkultúra területén”. A negyedik R. E. Dryer előadásban, amelynek címe „Legújabb megfigyelések az emberre patogén vírusok szövetkultúrában való viselkedéséről” volt, Enders az antibiotikumok bevezetését „kétségkívül a legfontosabb” tényezőnek nevezte, amely lehetővé tette a citopatogén hatások „vírusos” aktivitás bizonyítékaként való felhasználását. Az antibiotikumok valójában tökéletes ürügyet – vagy Enders szavaival élve „elvárást” – nyújtottak ahhoz, hogy a tenyészetek „vírusoktól” mentesek legyenek:
„Ma már olyan anyagokat lehet beoltani a szövetkultúrákba, mint torokváladék, széklet szuszpenzió vagy szervek, például a belek előkészítése, azzal az elvárással, hogy a legtöbb esetben nem fog nagyobb mikroorganizmusok növekedése bekövetkezni. Így felismerhetővé válik a vírus jelenléte, amely olyan megnyilvánulásokkal árulja el magát, amelyeket hamarosan leírok.”
E változás előtt a kutatóknak nem volt megbízható belső mutatójuk a „vírusok szaporodására” a szövetkultúrákban. Az egyetlen módja a „vírus” jelenlétének igazolására az volt, hogy állatokba oltottak be anyagot a kultúrákból, és megfigyelték a betegséget vagy a halált. Még Enders is elismerte ezt a korlátot:
„Egy másik helyzet, amely a múltban korlátozta a szövetkultúrák értékét a vírusok tanulmányozásában, az volt, hogy a kutatók nem tudtak a rendszerben megbízható indikátort találni a vírusok szaporodására. Ennek megfelelően a vírusok szaporodásának igazolásához a szövetkultúrából vett anyagot be kellett oltani egy fogékony állati gazdaszervezetbe. Gyakorlati szempontból tehát sok korábbi kutató számára a kultúra a legtöbb esetben nem kínált előnyt az állatokkal szemben a vírusok tenyésztése szempontjából. Sőt, sokkal kevésbé tűnt alkalmasnak, mivel a tenyészet nem adott egyértelmű fertőzésjeleket, amelyek összehasonlíthatók lennének a halállal vagy a kóros változások egyértelmű megjelenésével, amelyek általában kísérik a kórokozók in vivo szaporodását.”
A modern virológiai tankönyvek pontosan ugyanezt a korlátozást ismerik el. A Human Virology (4. kiadás) megjegyzi, hogy a korai szövettenyésztési munkák antibiotikumok nélkül szinte lehetetlenek voltak:
„De az igazán nagy áttörést az antibiotikumok felfedezése jelentette az 1940-es és 1950-es években. Addig nagyon nehéz volt a sejt- és szövetkultúrákat megóvni a levegőben terjedő baktériumok és penészgombák szennyeződésétől, de az antibiotikumok hozzáadása a tenyésztőközeghez gátolta ezeket a nemkívánatos szennyeződéseket, és lehetővé tette a kultúrák nyílt asztalon történő kezelését.”
Ez a beismerés döntő jelentőségű. Azt jelenti, hogy az a rendszer, amelyben a CPE a „vírusreplikáció” „bizonyítékává” vált, nem tudott működni a környezet mesterséges módosítása nélkül – a baktériumok elnyomásával, a természetes sejtviselkedés megváltoztatásával és olyan körülmények megteremtésével, amelyeknek nincs párja az élő szervezetekben. A „vírusreplikáció” „bizonyítéka” elválaszthatatlan volt a létrehozásához szükséges mesterséges rendszertől.
Az antibiotikumok bevezetésével Enders és mások most már feltételezhették, hogy ezek a tenyészetek hatékonyan „tiszták” a versengő organizmusoktól. Ez a technikai változás lehetővé tette, hogy ugyanazokat a degeneratív sejt hatásokat – amelyeket korábban szennyeződésnek vagy műterméknek tekintettek – „vírusos” aktivitás bizonyítékaként osztályozzák át. Az értelmezési keret nem azért változott, mert tisztított kórokozót mutattak ki, hanem azért, mert a kísérleti környezetet újradefiniálták.
Szerkezetileg Enders kijelentései és ellentmondásai azt mutatják, hogy a CPE „vírusos” replikáció bizonyítékaként való újraértelmezéséhez használt érvelés klasszikus logikai tévedéseket követett: a következmény megerősítését – feltételezve, hogy mivel CPE jelentkezett, „vírusos replikáció” is zajlik – és a kérdés elkerülését, azaz a CPE „immun” szérum általi gátlásának értelmezését annak bizonyítékaként, hogy „vírus” okozta azt. Tiszta és izolált „vírusok” nélkül, amelyek valódi független változóként szolgálhattak volna, a CPE-t a „vírusreplikációval” összekötő alapvető feltételezés soha nem került empirikus vizsgálatra.
Ennek ellenére 1954-ben Enders pontosan ezért kapta meg a Nobel-díjat. A kártyavár már megingott – éppen akkor, amikor a modern virológia alapjaiba építették.
Kísérleti kihívás: Ackermann 1954–1959
Érdekes módon, azt az alapfeltevést, hogy a CPE megfigyelése a tenyészetekben egyenlő a „vírusreplikációval”, pont abban az évben kérdőjelezték meg, amikor Enders és munkatársai díjat kaptak a feltevés „bizonyításáért”. 1954-ben Weston Wilbur Ackermann virológus, aki a 20. század közepén aktívan foglalkozott a polio kutatásával, publikálta a „A poliomyelitis vírus növekedési jellemzői HeLa sejtkultúrákban: a sejtkárosodás és a vírus szaporodása közötti párhuzamosság hiánya” című tanulmányát. (Wikipédia: A HeLa sejtek az első halhatatlan emberi sejtvonal, melyek Henrietta Lacks méhnyakrákos betegéből származnak, és 1951-ben vettek le róla orvosi vizsgálatok során anélkül, hogy ehhez hozzájárult volna. A sejtek egyedülállóan szaporodnak laboratóriumi körülmények között, forradalmasítva az orvostudományi kutatásokat, a vakcinák (pl. Polio) fejlesztésétől a súlytalansági kísérletekig. Ez az egyik legrégebbi és leggyakrabban használt emberi sejtvonal a tudományban.) Ebben Ackermann azt a kérdést tette fel, hogy a látható citopatológia elsődleges patológiának vagy „vírusos” szintézisnek és replikációnak tulajdonítható-e:
„Két lehetőséget vettünk figyelembe. Az egyik, hogy a fokozatosan fejlődő látható citopatológia a vírusanyag fokozatos növekedésével járó, fokozatosan fejlődő károsodás eredménye. A másik, hogy a lényeges károsodás a fertőzés kezdetével együtt befejeződik, amely esetben a látható patológia a vírus szaporodásától függetlenül alakulhat ki.”
Ackermann kifejezetten felállította azt a hipotézist, hogy a CPE párhuzamos lehet a „vírusos” replikációval, vagy nem, és a kísérleteket a klasszikus feltételezés, miszerint CPE = „vírusos” növekedés, közvetlen tesztjeként fogalmazta meg. Kísérletei során megfigyelte, hogy a sejtek pusztulása nyilvánvaló „vírusos” növekedés nélkül történt:
„A poliomyelitis vírus által indukált HeLa sejtek patológiája olyan metabolikus körülmények között zajlott, amelyek gátolják a vírusok szaporodását. Valószínűnek tűnik, hogy a két folyamat egymástól függetlenül működik, és a vírus szaporodását a relatív arányok határozzák meg.”
Ackermann a „vírusos” szintézist blokkoló módszereket alkalmazott, hogy tovább vizsgálja kérdéseit. Ehhez a fluoro-fenilalaninhoz (FPA) fordult, egy szintetikus aminosav-analóghoz, amely helyettesítheti a fenilalanint a fehérjeszintézisben. Amikor beépül a fehérjékbe, általában megzavarja azok térbeli szerkezetét és működését. Az akkori virológiai kutatásokban elméletileg a „vírusos” replikáció gátlására használták, mert a „vírusos” fehérjék deformálódnának vagy működésképtelenné válnának.
Ackermann HeLa sejteket tett ki az FPA hatásának, és megállapította, hogy bár az citosztatikus hatást fejtett ki (megállította a sejtek szaporodását), nem ölte meg a sejteket. Megerősítette, hogy az FPA nem inaktiválta közvetlenül a „poliovírust”, de amikor a sejteket annak jelenlétében „megfertőzték”, a „fertőző” titer szokásos emelkedése blokkolódott. Ennek ellenére a „fertőzött” sejtek továbbra is citopátiás változásokat mutattak, és a szokásos ütemben degenerálódtak. Nagy dózisú „fertőzések” esetén szinte az összes sejt normálisan elpusztult, de alacsony dózisú beoltások esetén csak néhány sejtnél jelentkeztek citopátiás hatások, és a károsodás nem terjedt át a szomszédos sejtekre. Normális esetben a „fertőzés” terjedt volna a tenyészetben, de ebben az esetben korlátozott maradt, ami arra utalt, hogy új „vírus” nem termelődött és nem terjedt tovább.
Ebből Ackermann arra a következtetésre jutott, hogy a „vírusreplikáció” és a sejtkárosodás elkülöníthető jelenségek, azaz a CPE nem függött szigorúan a „vírus szaporodásától”. A folyamatokat „jelentős fokú autonómiával” jellemezte. Más szavakkal, a kísérlet arra utalt, hogy bármi is okozza a sejtkárosodást (amit a virológusok „vírusos citopatogenitásnak” értelmeztek), az nem egyszerűen azonos a „vírusreplikációval”. Ez az eredmény aláásta azt a feltételezést, hogy a tenyészetben bekövetkező sejthalál = „vírusreplikáció” = „vírusaktivitás” bizonyítéka:
„Bár a fluor-származék teljesen gátolta a vírus szaporodását, nem akadályozta meg a vírus citopatogén hatását. Fluor-fenilalanin jelenlétében a fertőzött sejtek szétesése a szokásosnak tűnő sebességgel folyt, a fertőző ágens növekedése nélkül. Kísérletileg kimutatták, hogy a vírus szaporodásához és a sejtek károsodásához vezető folyamatok jelentős fokú autonómiával rendelkeznek.”
Ez közvetlen kísérleti cáfolata annak a feltételezésnek, hogy a CPE „vírusreplikációt” jelezne a saját szabványok és módszertan alkalmazásával.
Ackermann a citoplazmatikus granulátumokat és a sejtmagban detektált változásokat is a HeLa sejtekben előforduló „poliovírus-fertőzés” „feltűnő” és következetes jellemzőjeként írta le, azt állítva, hogy ezek a kontrollcsoportban nem voltak jelen. Ugyanakkor elismerte, hogy ezek néha kémiai sérülés vagy „toxikus” széklet szuszpenzióknak való kitettség után jelentek meg. Ez az elismerés megcáfolja a specifikusságra vonatkozó állítást: ha ugyanazok a morfológiai hatások „nem vírusos” károsodásokból is származhatnak, akkor azok nem szolgálhatnak egyedi „vírusos” ok bizonyítékaként. Legjobb esetben is csak egy nem specifikus sejtes stresszreakciót mutatnak, amelyet tévesen „vírusos” aktivitás bizonyítékaként értelmeznek:
„(Ackermann 1954-es tanulmányának) 4. ábráján a fertőzés után 21 órával készült tenyészet látható. A sejtek többsége már nem tapad a fedőüveghez. Azok a kevesek, amelyek tapadnak, jelentős sejtmag piknózist (a sejtmag anyagának összesűrűsödése erősen festődő tömeggé) mutatnak, és feltűnő eredmény, hogy sok mélykékre festődő citoplazmatikus granulátum található bennük, amelyek mérete és alakja szabálytalan. Ezek következetesen megtalálhatók a HeLa sejtekben, amelyek citopatológiás vizsgálatnak vannak alávetve a I. és III. típusú poliomyelitis vírus számos törzsével történő fertőzés után. Ezek nem figyelhetők meg kontrollkultúrákban, és csak ritkán fordulnak elő olyan kultúrákban, amelyeket hosszú ideig inkubáltak táptalajcsere nélkül, vagy olyan kultúrákban, amelyeket különböző kémiai anyagok vagy „toxikus” széklet szuszpenziók károsítottak.”
Ackermann eredményei azt mutatják, hogy a tenyészetben a sejtek elhalása függetlenül a „vírusreplikációtól” is bekövetkezhet, és toxinok vagy kémiai sérülések is kiválthatják. A CPE messze nem erősíti meg a „vírusos” okot, hanem inkább egy nem specifikus stresszreakciónak tekinthető, amelyet tévesen „vírusos” aktivitás bizonyítékaként értelmeznek.
Egy 1958-as, „A poliovírus citopatogén hatása; bizonyíték egy extravirális toxinra” című követő tanulmányban Ackermann megerősítette ezeket a megállapításokat, kimutatva, hogy a sejtkárosodás és a „vírus” termelés autonóm folyamatok, közvetlenül cáfolva azt a feltételezést, hogy a CPE bizonyítja a „vírusreplikációt”. Sajnos a cikk fizetős, de a rendelkezésre álló összefoglalóból némi képet kaphatunk a munkáról:
„Az 1-es típusú poliovírussal fertőzött szövetkultúrákból nyert folyadékok toxikus hatását figyelték meg, amikor a folyadékokat friss kultúrákba juttatták. A destruktív aktivitás abban különbözik a szokásos vírus által kiváltott citopatogén hatástól, hogy gyorsan jelentkezik, hosszan tartó lappangási időszak és vírustermelés nélkül. A fertőző aktivitás szelektíven semlegesíthető antiszérummal, a toxicitás teljes megőrzése mellett. Bizonyos antiszérumok azonban a toxint is semlegesítik. Ezenkívül a két aktivitás hőstabilitása eltérő. A sejtek elpusztítására és kimutatható mennyiségű toxin termelésére való képesség csak a poliovírus bizonyos törzseinek jellemzője.”
Szerencsére Ackermann munkájáról további részletek találhatók a Cabi Digital Library-ben. A kutatók leírták, hogy a „poliovírussal fertőzött” tenyészetekben a sejtek elvesztése két különálló fázisban történt: egy korai sejtpusztulási hullám 2–4 órán belül, mielőtt bármilyen kimutatható „vírus” megjelent volna a sejtekben, és egy későbbi hullám 6–12 óra múlva, egybeesve a feltételezett „intracelluláris vírus” megjelenésével. A korai pusztulást a tenyészetfolyadékban jelen lévő toxin-szerű tényezőnek tulajdonították. Döntő fontosságú, hogy a toxikus aktivitás eltérően viselkedett a „fertőzőképességtől”: míg a „vírusfertőzőképesség” 4 °C-on hetekig stabil maradt, a toxicitás gyorsan csökkent, ami azt mutatta, hogy ezek elkülöníthető jelenségek. Az antiszérumkísérletek megerősítették ezt a különbséget: egyes szérumok semlegesítették a „fertőzőképességet”, de nem a toxicitást, míg mások mindkettőt blokkolták, amíg HeLa sejtekkel nem adszorbeáltak, ami után csak a „fertőzőképesség” gátlódott.
Ezek az eredmények azt mutatják, hogy a CPE nem a „vírus szaporodásának” elkerülhetetlen következménye volt, hanem instabil, sejtekből származó toxikus melléktermékekből származhatott. Ackermann eredményei messze nem bizonyították a „vírusreplikációt”, hanem a tenyésztésen alapuló vizsgálatok kétértelműségét tárták fel, és arra utaltak, hogy amit a virológusok „citopatogenitásnak” neveztek, gyakran a kísérleti rendszerük nem specifikus toxikus hatása volt:
„Számos vírus esetében írtak le a fertőzőképességtől független toxikus hatást. Ez a cikk olyan kísérleteket ismertet, amelyek arra utalnak, hogy a poliovírussal fertőzött szövetkultúrák toxint és fertőző vírust is tartalmaznak:
1. A nagy dózisú poliovírussal (1-es típusú Mahoney PT törzs) fertőzött tenyészetből a sejtek elvesztése két szakaszban történik: –
(a) a fertőzés után 2-4 órával, az intracelluláris vírus megjelenése előtt;
(b) 6-12 órával, az intracelluláris vírus megjelenése után. Az első időszakot a vírus toxinjával hozzák összefüggésbe.2. A poliovírus 4 °C-on több hétig történő tárolása esetén a fertőzőképesség alig vagy egyáltalán nem csökken, de a toxikus aktivitás gyorsan csökken.
3. Olyan antiszérumok állíthatók elő, amelyek csak a vírus fertőzőképességét semlegesítik, a toxicitását nem. Így egy majom antiszérum a Brunhilde 1-es típusú vírust semlegesítette, de a toxicitását nem. Hasonlóképpen, a majmokból előállított, 1-es típusú vírussal fertőzött HeLa sejtkultúrák folyadékából készült antiszérum úgy a fertőzőképességet, mint a toxicitást gátolta, de a HeLa sejtekkel való adszorpció után csak a fertőzőképességet befolyásolta. A toxicitás elleni hatás valószínűleg nem a HeLa sejtek ellen irányuló antitesteknek volt köszönhető, mivel
(a) egy másik, HeLa sejtek ellen irányuló antiszérum nem volt hatékony,
(b) a HeLa sejtek kivonatai nem voltak toxikusak.
4. Úgy tűnik, hogy nem minden poliovírus-törzs termel toxint. Így a Mahoney 1-es típusú poliovírus PT törzse (HeLa sejtekben átvitt, nagy plakkot képező) toxint termelt HeLa és majom vese sejtekben, míg a szintén Mahoney 1-es típusú pt törzs (majomban és majom vese sejtekben átvitt, kis plakkot képező) egyik sejtkultúrában sem termelt toxint.
Ackermann eredményei pontosan azok, amikre számítani lehet, ha a CPE-t stresszes vagy haldokló sejtek instabil, toxikus melléktermékei okozzák, és nem egy replikáló mikroba. A toxin instabilitása a „fertőzőképességhez” képest, a törzsek és átmenetek közötti variabilitása, valamint az antiszérumok általi szelektív semlegesítése mind azt erősítik, hogy ez nem specifikus sejttoxicitás volt. Ackermann munkája ahelyett, hogy megerősítette volna a CPE-t a „vírusreplikáció” indikátoraként, feltárta a virológia alapjainak végzetes hibáját: a CPE nem bizonyíték a „vírus” létezésére. IDÁIG
Az 1959-es, „Vírussal fertőzött sejtek biokémiai vizsgálatai” című cikkében Ackermann tovább gyengítette a „vírusreplikáció” narratíváját azzal, hogy rámutatott: az úgynevezett „vírussal fertőzött” sejtekben termelődő új anyagok többsége egyáltalán nem volt „vírusos”. A puszta mennyiség, a szubcelluláris eloszlás és az RNS-összetétel alapján megjegyezte, hogy a „poliovírus-fertőzés” során kialakult termékek nagy része egyértelműen sejtes természetű volt, és hasonló eredményeket jelentettek a „herpeszvírus” és az „adenovírus” esetében is. A biokémiai vizsgálatok valójában több glikolitikus enzim – normális metabolikus fehérje – emelkedett szintjét mutatták ki ugyanabban a citoplazmatikus frakcióban, ahol a fehérjetartalom növekedését mérték. Ezek a megfigyelések ismét arra utaltak, hogy amit a virológusok „virális szintézisként” értelmeztek, az inkább nem specifikus sejtes stresszreakció vagy metabolikus zavar volt, nem pedig új „vírusok” keletkezése.
„A kísérleti bizonyítékokból (a részt vevő anyagok mennyisége, azok eloszlása a sejt alkatrészei között és az RNS bázisösszetétele) egyértelműen kitűnik, hogy a poliovírus-fertőzés során kialakult anyagok nagy része valóban nem vírus típusú (Maassab et al., 1957; Loh et al., 1958). Kizárólag mennyiségi szempontok alapján úgy tűnik, hogy hasonló eredmények születnek a herpeszvírus (Newton és Stoker, 1958) és az adenovírus (Boyer et al., 1957) fertőzés esetén is. A második pont, az újonnan képződött anyag sejtes természetére vonatkozóan, további vizsgálatot igényel. A poliovírus esetében azonban különösen érdekes a fertőzés után megemelkedett hat glikolitikus enzimszintről szóló legújabb jelentés (Matzelt et al., 1958), ha figyelembe vesszük, hogy ezek normális esetben a citoplazma oldható frakciójában találhatók. Ez megfelel a 111-es frakciónak, amelynek teljes fehérjetartalmát növekedni találtuk (Loh et al., 1958).”
Ackermann elismerte, hogy a CPE egyáltalán nem „vírus” okozta lehet, hanem a sejt saját kiegyensúlyozatlan növekedése vagy a tenyészetben felhalmozódó toxikus melléktermékek. Más szavakkal, a virológusok által használt jelző nem specifikus és nem különböztethető meg a kémiai károsodástól:
„Ebből a gondolatmenetből adódik a nehézség, amikor a vírus által stimulált normális aktivitás nem egyezik a sejt egyéb normális tevékenységeivel. A citopatogén hatás elsődleges oka nem a kis mennyiségű idegen anyag szintézise, hanem a sejt kiegyensúlyozatlan növekedése lehet. A morfológiai integritás végleges elvesztése azonban bizonyos anyagok másodlagos felhalmozódásának eredménye lehet. Egy ilyen citotoxikus anyagot egy előzetes publikációban már leírtak (Ackermann et al., 1958~).”
Összességében Ackermann munkája a virológia saját módszereinek és szabványainak felhasználásával kimutatta, hogy a CPE nem megbízható mutatója a „vírusreplikációnak”. Ehelyett ugyanolyan könnyen magyarázható a sejtek folyamatainak zavarával és a tenyésztési rendszer toxikus melléktermékeivel. Kritikus megállapításai – különösen a toxin által kiváltott citopátiás hatás azonosítása – tartós hatással voltak arra, ahogyan a virológusok értelmezik a tenyésztésen alapuló vizsgálatokat és a „vírusfertőzés” biokémiai alapjait. Döntő fontosságú, hogy feltárták, mennyire nem specifikus és félrevezető volt valójában a tenyésztésen alapuló vizsgálat.
A falszifikálhatatlan érvelés megjelenése [3] 
Ezeket a kísérleti kihívásokat követően világossá vált, hogy a CPE előfordulhat kimutatható „vírusreplikáció” hiányában is, és ahogy Enders maga is elismerte, a „vírusreplikáció” megfigyelhető CPE nélkül is előfordulhat. Ezért a virológia egy sor értelmező „mentőeszközt” fejlesztett ki a domináns hipotézis megőrzése érdekében. Amikor a citopátiás hatások (CPE) nem feleltek meg az elvárásoknak, új kategóriákat hoztak létre, mint például a „nem citocidális fertőzés”, „perzisztens fertőzés”, „abortív fertőzés” vagy „gazdaszervezet-sejt variabilitás”. Ahogy H.G. Pereira virológus „Az állati vírusok citopátiás hatása” című művében elismerte, még a CPE és a „vírusreplikáció” közötti alapvető kapcsolat is inkonzisztens volt:
„A citopátiás hatás a vírusfertőzőképesség gyakori megnyilvánulása, és általában teljesen fertőző vírusrészecskék vagy nem fertőző vírusanyagok szintézisével jár együtt. Azonban számos rendszerben megfigyelték, hogy CPE jelentkezik vírusanyagok kimutatható termelődése nélkül, és fordítva, vírusmultiplikáció jelentkezik nyilvánvaló CPE nélkül.”
Ez a megállapítás megdöbbentő volt: a CPE előfordulhatott anélkül, hogy bármilyen „vírus” kimutatható lett volna, és a „vírus szaporodása” állítólag látható citopátiás hatás nélkül is bekövetkezhetett. Mindkét esetben az ok-okozati összefüggés inkább újraértelmezésen, mint empirikus megerősítésen alapult.
Még ennél is árulkodóbb, hogy Pereira dokumentálta, hogy az úgynevezett „vírusos CPE” nem volt állandó, önálló jelenség, hanem egyszerűen a tenyésztési körülmények megváltoztatásával indukálható vagy elnyomható volt. Például Bang és munkatársai (1957) arról számoltak be, hogy a keleti ló-encephalomyelitis „vírussal” való feltételezett „fertőzésből” való gyógyulás akkor volt valószínűbb, ha embrió-kivonatot adtak a táptalajhoz, és az inkubáció 37 °C-on történt, míg a krónikus „fertőzés” fenntartásához alacsonyabb hőmérséklet és ugyanaz az embrió-kivonat volt szükséges. Hasonlóképpen, Ginsberg (1958) és Takemoto & Habel (1959) kimutatták, hogy a hiányos táptalajok használata a CPE növekedéséhez vezetett, amit aztán a „vírustermelés” növekedésével azonosítottak. Pereira továbbá megjegyezte, hogy az elsődleges szövetkultúrák egyszerűen az öregedésükkel „fokozatosan növekvő érzékenységet mutattak a vírusos CPE-re” (Kumazai et al., 1958; Frothingham, 1959; Chaproniere, 1960). Más szavakkal, a CPE jelenléte, hiánya vagy súlyossága a tápanyagok rendelkezésre állása, az inkubációs hőmérséklet, az adalékanyagok vagy a tenyészet kora által befolyásolható – ezek a tényezők teljesen függetlenek bármely állítólagos „vírustól”.
Pereira végül következtetésében elismerte, hogy egyetlen CPE sem alkalmas egyedülállóan az úgynevezett „vírusok” diagnosztizálására:
„A főbb víruscsoportok CPE-jének jellemzésére tett kísérletet az idézett mű I. táblázata tartalmazza. Hangsúlyozni kell, hogy a táblázatban felsorolt jellemzők egyike sem patognomonikus egy adott vírus által okozott fertőzésre, és hogy egyes vírusok bizonyos körülmények között kivételes viselkedést mutathatnak. Ha azonban ezeket a jellemzőket a citopátiás hatás összképének részeként tekintjük, akkor értékes bizonyítékot nyújtanak, amely sugallja vagy alátámasztja a különböző vírusok közötti lehetséges kapcsolatok létezését.”
Ez egy rendkívüli beismerés. Nemcsak hogy egyetlen citopátiás hatás sem patognomonikus (azaz önmagában a diagnózist megalapozó jel), hanem mindig vannak kivételek, és az eredmények csak „utalhatnak” kapcsolatokra, de nem bizonyíthatják az ok-okozati összefüggést. Más szavakkal, azt a jelenséget, amelyet egykor a „vírusreplikáció” határozott bizonyítékaként kezelték, most nem specifikusnak, inkonzisztensnek és újraértelmezhetőnek ismertek el.
A modern virológiai tankönyvek is visszhangozzák ezeket a beismeréseket. A Human Virology (4. kiadás) nyíltan leírja azokat a „vírusokat”, amelyek nem okoznak látható CPE-t, csak közvetett módszerekkel, például „szuperfertőzés-rezisztenciával” vagy immunfluoreszcenciával mutathatók ki, és egyes esetekben egyáltalán nem tenyészthetők sejtkultúra-rendszerekben.
„Néhány vírus, bár szaporodik a sejtkultúrában, nem okoz látható CPE-t, és csak azon képességük alapján mutatható ki, hogy a sejteket rezisztenssé teszik egy második vírus szuperinfekciójával szemben. Más, CPE-t nem okozó vírusok immunfluoreszcenciával (36.1. ábra) vagy vörösvérsejtekhez való kötődési képességükkel (haemadsorption) mutathatók ki. Egyes vírusok, pl. bizonyos bél- és hepatitis vírusok nem tenyészthetők sejtkultúra-rendszerekben.”
A szerzők még azt is megjegyzik, hogy a „HIV izolálásához” mesterségesen kellett stimulálni a limfocitákat növényi lektinekkel és IL-2-vel, mert a limfociták „normális körülmények között nem tarthatók kultúrában”. Más szavakkal, még a mainstream virológia szerint is, a jelenség, amelyet egykor a „vírusreplikáció” elsődleges bizonyítékaként kezeltek, soha nem volt univerzális, megbízható vagy önmagát igazoló.
Az Amerikai Mikrobiológiai Társaság a Cytopathic Effects of Viruses Protocols (Vírusok citopátiás hatásai protokollok) című dokumentumában elismerte, hogy sok „vírus” egyáltalán nem okoz CPE-t az állítólagos természetes gazdasejtjeiben:
„Egyes vírusok egyáltalán semmi vagy csak minimális okoznak CPE-t okoznak természetes gazdasejtjeikben. Jelenlétük vizuálisan csak hemadszorpcióval [4] vagy interferenciával mutatható ki, amelynek során a CPE-t nem mutató fertőzött sejtkultúrák gátolják a kultúrákba később bejuttatott másik vírus replikációját, vagy in situ vírusantigénnel vagy nukleinsav-detektálással.”
Arra is figyelmeztetnek, hogy nincs rögzített szabvány arra nézve, hogy a CPE-nek hogyan kell kinéznie:
„Ne feledje, hogy egy adott vírus nem feltétlenül felel meg a családjára vonatkozó normának, vagy különböző gazdasejt-típusokban különböző CPE-t okozhat.”
CPE hiányában közvetett kimutatási módszereket alkalmaztak, mint például hemadszorpció, interferencia-vizsgálatok és molekuláris technikák. Cann Molekuláris virológia alapjai című műve kifejezetten elismeri a CPE történelmi szerepét a replikáció helyettesítőjeként, valamint azt, hogy ha ez nem fordul elő, akkor végül a genetikai elemzésre kell támaszkodni:
„Azoknál a vírusoknál, amelyeknél ilyen rendszer nem létezik – mert nem citopátiásak, nem replikálódnak tenyészetben, vagy nem okoznak helyi elváltozásokat –, a molekuláris genetika kialakulása előtt alig volt lehetséges genetikai elemzés, utána azonban sok esetben rendkívül gyors volt a fejlődés.”
Hasonlóképpen, a Fields Virology is elismerte ugyanezt a pontot – ha a CPE nem figyelhető meg, más módszereket kell alkalmazni:
„A citopátiás hatás a fertőzés legegyszerűbb és legszélesebb körben alkalmazott kritériuma, de nem minden vírus okoz citopátiás hatást, és ezekben az esetekben más módszerekkel kell beérni.”
Továbbá elismerte, hogy a „vírusok” magas titerig növekedhetnek anélkül, hogy CPE-t okoznának, ezért más detektálási módszerekre van szükség:
„A vírusok sejtkultúrában történő szaporodását leggyakrabban a mikroszkóposan látható citopátiás hatás (CPE) kialakulása alapján észlelik. Egyes vírusok azonban magas titerig szaporodhatnak anélkül, hogy látható CPE-t okoznának, ezért más módszerekkel kell kimutatni őket.”
Ez nem csupán elméleti elismerés volt, hanem a mindennapi laboratóriumi gyakorlatot is meghatározta. Például Minamoto és munkatársai (1976) leírják, hogy a a veszettség „vírusának” növekedése meghatározására hemadszorpciót használták helyettesítő indikátorként:
„A HAD jelenséget a veszettség SN-tesztjében alkalmazták a vírus szaporodásának indikátoraként a citopátiás hatás (CPE) helyett.” (Journal of Virology, 20(1): 234–241).
Más szavakkal, a citopátiás destrukció hiánya soha nem cáfolta a hipotézist. Ehelyett a bizonyítékok hálóját bővítették: minden megfigyelhető változás „vírusreplikációként” értelmezhető volt, a CPE hiányát pedig alternatív mechanizmusok vagy közvetett molekuláris helyettesítők segítségével magyarázták. Az idő múlásával a hangsúly a közvetlen, látható hatásokról (amelyek már eleve kétértelműek és mesterségesek voltak) a biokémiai nyomokra és genetikai fragmensekre helyeződött át, amelyek maguk is előfeltételezik azokat a jelenségeket, amelyeket bizonyítaniuk kellene.
Ezek a stratégiák a hipotézist cáfolhatatlanná tették: bármely CPE-mintázat utólagosan „replikáció” bizonyítékaként értelmezhető volt, és a CPE hiánya alternatív mechanizmusokra vagy közvetett helyettesítőkre hivatkozva magyarázható volt. Ezzel a logikával egyetlen lehetséges eredmény sem számíthatott a „vírus” létezését vagy aktivitását cáfoló bizonyítéknak. Az eredmény az volt, hogy a virológia hatékonyan elszigetelte alapvető feltételezését az empirikus cáfolattól.
Logikai buktatók és cáfolhatatlan megerősítés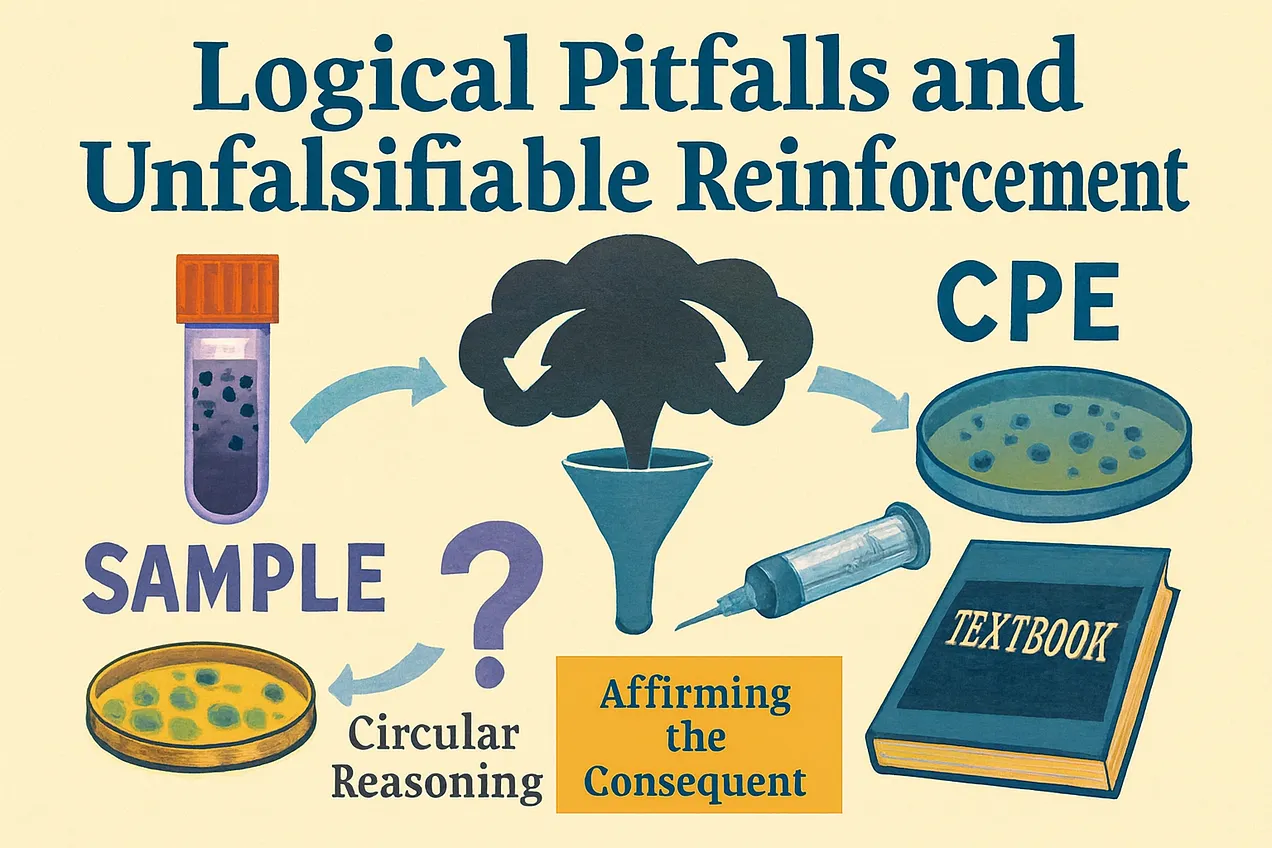
A CPE-érvelés alakulása jól illusztrálja, hogyan gabalyodott bele a klasszikus virológia a logikai buktatók és értelmezési menekülési utak hálójába. Bár Enders előtt is megfigyelték a citopátiás hatásokat, ezeket a megállapításokat többnyire szennyeződésnek, technikai hibának vagy laboratóriumi artefaktumnak tekintették. Nem a jelenség maga változott meg, hanem az értelmezési környezet. Az antibiotikumok megjelenésével – és a poliomyelitis elleni küzdelemre irányuló intenzív társadalmi és politikai nyomás alatt – Enders képes volt ezt a korábban figyelmen kívül hagyott laboratóriumi hatást „vírus” jelenlétének jelentőségteljes biológiai bizonyítékaként újraértelmezni. A korábban zajként kezelt jelenséget bizonyítékként használták fel.
Ettől kezdve a citopátiás hatást (CPE) a „vírusreplikáció” bizonyítékaként kezelték, annak ellenére, hogy ezt a következtetést soha nem igazolták tisztított „vírus” készítmények felhasználásával. Az érvelés klasszikus logikai tévedéseken alapult. Mivel a „vírusok” feltételezett sejtkárosító hatása miatt a sejtkárosodás megfigyelését a „vírus” replikációjának bizonyítékaként értékelték – ez egy példa a következmény megerősítésére.
Ezt a körkörösséget az „immun” szérum használata erősítette meg: amikor az „antitestek” gátolták a CPE-t, ezt bizonyítékként kezelték arra, hogy „vírus” okozta, annak ellenére, hogy az ok-okozati láncot soha nem állapították meg függetlenül. Amikor a CPE nem jelent meg, az alapfeltevés változatlan maradt, és alternatív „vírusos” magyarázatokat vezettek be annak megőrzése érdekében. Minden esetben a „vírus” létezését és aktivitását feltételezték, ahelyett, hogy bizonyították volna.
Ahogy a ellentmondások növekedtek, a kapufélfák eltolódtak. A CPE-t kezdetben meghatározó kritériumként kezelték, de amikor a kivételek száma növekedett, a „vírusreplikáció” definícióját kiterjesztették a hemadszorpcióra, az interferenciára, az antigénfestésre vagy a genetikai fragmensekre. A végeredménytől függetlenül mindig azt állították, hogy „replikáció” történik.
Az eredmény egy olyan keretrendszer lett, amelynek célja a falszifikáció elleni védekezés volt. Ahhoz, hogy egy hipotézis tudományos legyen, léteznie kell olyan megfigyelésnek, amely bizonyíthatja, hogy téves. Itt azonban mind a CPE jelenléte, mind hiánya, valamint bármely közvetett biokémiai marker a replikáció bizonyítékaként lett újraértelmezve. A felépítés miatt a feltételezést nem lehetett cáfolni, csak megerősíteni.
A CPE története egy nagyobb tanulságot is hordoz arról, hogy a tudomány hogyan térhet el a helyes útról, ha a feltételezéseket nem tesztelik, hanem védik. Ezt a hamisíthatatlan struktúrát tekintélyes díjak és tankönyvek is megerősítették, biztosítva, hogy a kutatók generációi örököljék a feltételezést, ahelyett, hogy megkérdőjeleznék. A CPE esete megmutatja, milyen könnyen téveszthető össze a korreláció az ok-okozati összefüggéssel, és hogyan válhatnak a logikai tévedések intézményesített tudományos módszerekké. A CPE-re való történelmi támaszkodás, amelyet utólagos mentőeszközök is megerősítettek, megerősítette a megcáfolhatatlan doktrínát, amelyet megalapozatlan érvelés támasztott alá. Ez aláhúzza a független ellenőrzés és a valódi megcáfolhatóság szükségességét a tudományban, megmutatva, hogy egy nem tesztelt feltételezés hogyan válhat dogmává és alakíthatja az évtizedek gyakorlatát.
A virológia nem volt képes szembenézni hibás alapvető érvelésével, és figyelmen kívül hagyta azt az elvet, amelyet az igazi tudomány megkövetel: a hipotéziseknek tesztelhetőnek kell lenniük, és lehetségesnek kell lennie, hogy tévesnek bizonyuljanak. A kezdetektől fogva ez a tudományág a bizonyítást feltételezésekkel helyettesítette. A hatásokat okként kezelték. A hiányzókat jelenlétként ábrázolták. A körkörös logikát pedig módszertani szabványnak emelték.
Ebben a fényben a citopátiás hatások beállítása „a vírusok” létezésének és replikációjának bizonyítékaképp nem a tudomány diadala, hanem annak hiányát jelzi. A virológia nem távolodott el az empirikus szigorúságtól – eleve anélkül épült fel.
Hivatkozások:
[1] Például itt: Az általánosan elfogadott víruselmélet cáfolata – Alapok
[2] Az inkluziós testek sűrű, aggregált fehérje- vagy egyéb anyagok (pl. vírusok, zsírok, glikogén) halmazai, amelyek a sejtek belsejében (magban vagy citoplazmában) találhatók, és gyakran stresszt, vírusfertőzést vagy genetikai rendellenességeket jeleznek, sejtek „tárolóhelyeként” vagy vírusreplikációs helyként működnek, és olyan betegségekben jelennek meg, mint a Parkinson-kór, a veszettség és az inkluziós testes myositis. Akkor alakulnak ki, amikor a sejtek túltermelnek fehérjéket, vagy nem tudják azokat megfelelően feldolgozni, így oldhatatlan csomók keletkeznek, amelyek zavarhatják a sejtek működését.
[3] Karl Popper szerint egy elmélet, hipotézis akkor tudományos, ha bizonyítható és cáfolható (verifikálható és falszifikálható), azaz alávethető bizonyítási és cáfolási eljárásnak.
[4] A hemadszorpció egy biológiai jelenség, leggyakrabban a virológiában használják vírusok kimutatására, ahol a fertőzött sejtek felületén megjelenő vírusfehérjék (pl. hemagglutinin) képesek vörösvértesteket (VVT-ket) megkötni, hidakat képezve a sejtek és a VVT-k között.
2025. december
Közzéteszi:
Király József
